ОБМІН РЕЧОВИН У КЛІТИНІ
ОБМІН РЕЧОВИН У КЛІТИНІ
| Сайт: | Підготовка до ЗНО - Освітній портал "Академія" |
| Курс: | Підготовка до ЗНО з біології |
| Книга: | ОБМІН РЕЧОВИН У КЛІТИНІ |
| Надруковано: | Гість |
| Дата: | Thursday 3 July 2025 2:15 PM |
1. ОБМІН РЕЧОВИН У КЛІТИНІ
ОБМІН РЕЧОВИН У КЛІТИНІ
Обмін речовин (метаболізм) – сукупність хімічних перетворень, які відбуваються в клітинах та забезпечують їхній ріст, життєдіяльність і відтворення.
Обмін речовин живої клітини складається з двох протилежно направлених видів реакцій – катаболічних і анаболічних.
Катаболізм або енергетичний обмін – сукупність реакцій розпаду органічних сполук.
Анаболізм або пластичний обмін – сукупність реакцій синтезу органічних сполук
Під час розщеплення (катаболізму) органічних сполук (білків, жирів, вуглеводів) виділяється енергія, яка акумулюється в хімічних зв'язках молекул АТФ. Ця енергія використовується клітиною в анаболічних процесах – синтезі власних, необхідних на даний момент часу білків, жирів і вуглеводів. Таким чином, енергетичний і пластичний обмін тісно пов'язані між собою потоками речовини й енергії.
За способом добування енергії живі організми поділяються на автотрофів і гетеротрофів. Клітини гетеротрофів (тварини, гриби, більшість бактерій, паразитичні рослини) для побудови власних біополімерів використовують мономери поглинених і розщеплених ними органічних сполук, а також енергію, що виділяється при цьому. Автотрофи здатні синтезувати власні біополімери з води, вуглекислого газу та мінеральних солей. Залежно від джерела енергії, яка використовується, автотрофи поділяють на фототрофів (акумулюють сонячну енергію – зелені рослини, деякі бактерії) і хемотрофів (акумулюють енергію, що виділяється внаслідок окисно-відновних процесів,– сіркобактерії, нітрифікуючі бактерії).
За чутливістю до концентрації кисню в навколишньому середовищі всі організми поділяються на аеробів і анаеробів. Аероби можуть існувати тільки за наявності достатнього вмісту кисню. Анаероби поділяються на облігатних (існують тільки в безкисневих умовах) і факультативних (здатні до життя в широкому діапазоні концентрацій кисню).
ОБМІН ВУГЛЕВОДІВ
Енергетичний обмін вуглеводів
|
Етап |
Місце |
Процеси |
|
Підготовчий |
Шлунково-кишковий тракт, у цитоплазмі клітин |
Органічні макромолекули за участю ферментів розпадаються на дрібні молекули: |
|
Безкисневий (анаеробний, гліколіз, неповне розщеплення) |
На внутрішньоклітинних мембранах гіалоплазми |
Гліколіз: |
|
Кисневий (аеробний, тканинне дихання) |
У матриксі мітохондрій |
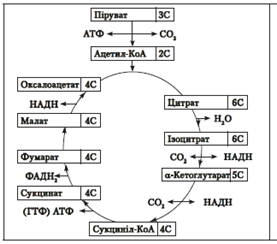
Цикл Кребса: суть перетворень полягає у ступінчастому декарбоксилюванні й дегідруванні піровиноградної кислоти, під час яких утворюються АТФ, НАДН і ФАДН2. |
|
Сумарне рівняння енергетичного обміну: C6H12O6 + 6O2 + 38АДФ + 38Ф – 6CO2 + 42H2O + 38АТФ |
||
Пластичний обмін вуглеводів у гетеротрофних організмів
Моносахариди, які потрапили в цитоплазму, можуть не тільки піддаватися розщепленню з виділенням енергії, але й є матеріалом для синтезу власних полісахаридів клітини.
Глюкоза за допомогою специфічних ферментів полімеризується з утворенням глікогену (цей процес називається глікогенезом). У цьому разі витрачається енергія АТФ. Синтезований глікоген накопичується в цитозолі у вигляді гранул і є запасною поживною речовиною. У разі необхідності він окиснюється до глюкози, яка включається в гліколіз. Утворена внаслідок розпаду глікогену в клітинах печінки ссавців глюкоза виходить у кров і є джерелом енергії для нейронів і м'язів. Оскільки більшість реакцій гліколізу є оборотними, клітина здатна синтезувати глюкозу з інших сполук ацетил-КоА, кислот, залучених у ЦТК (цей процес називається глюконеогенезом).
Пластичний обмін вуглеводів у фототрофних організмів – фотосинтез
Фотосинтез – це процес перетворення енергії сонячного світла на енергію хімічних зв'язків і синтезу органічних сполук (вуглеводів) з неорганічних (вода та вуглекислий газ).
Фотосинтез – процес утворення органічних сполук із неорганічних завдяки перетворенню світлової енергії в енергію хімічних зв'язків. Здійснюється в клітинах зелених рослин за участю пігментів хлоропластів – хлорофілів a та b (зелені), каротиноїдів (жовті), фікобілінів (сині та червоні).
|
Фази |
Місце |
Процеси |
|
Світлова фаза |
На мембранах тилакоїдів хлоропластів |
Фотосинтезуючі пігменти поглинають енергію світла, що приводить до «збудження» одного з електронів молекули пігменту, який за допомогою молекул-переносників переміщується на зовнішню поверхню мембрани тилакоїдів. Відбувається фотоліз води: H2O ^ H+ + OH-. Йони H+ перетворюються на Гідроген, який використовується у реакціях фотосинтезу: H+ + e → H . Гідроксильні йони, взаємодіючи між собою, утворюють кисень, воду й вільні електрони: 4OH → 2H2O + O2 + 4 e . Електрони через ряд проміжних речовин передають енергію для відновлення НАДФ (нікотинамідаденіндинуклеотидфосфат), який приєднує два атоми Гідрогену й перетворюється на НАДФ+ Частина енергії електронів перетворюється на енергію АТФ: АДФ + Ф + Q → АТФ |
|
Темнова фаза (світло не потрібне) |
У стромі хлоропластів |
За наявністю CO2, енергії АТФ та сполук, що утворилися у світлових реакціях, відбувається приєднання Гідрогену до CO2, який надходить до хлоропластів із зовнішнього середовища. Через ряд послідовних реакцій (цикл Кальвіна) за участю специфічних ферментів утворюються різноманітні сполуки, основними з яких є вуглеводи У процесі цикла Кальвіна відбувається фіксація атома Карбону CO2 для побудови глюкози (C6H12O6) з рибульозо-1,5- дифосфату (C5H8O5P2). Для синтезу 1 молекули глюкози в циклі Кальвіна необхідно 12 молекул НАДФН і 18 молекул АТФ, що утворюються в результаті фотохімічних реакцій фотосинтезу. Енергія для синтезу вуглеводів утворюється внаслідок розщеплення молекул АТФ, синтезованих під час проходження електронів по компонентах ФС1 і ФС2. Утворена в процесі циклу Кальвіна глюкоза може потім розщеплюватися до пирувата, надходити в цикл Кребса. |
|
Сумарне рівняння фотосинтезу: 6CO2 + 6H2O +ен е ргія → C6H12O6 + 602 |
||
Значення фотосинтезу.
Фотосинтез – єдиний процес у біосфері, що веде до збільшення її вільної енергії за рахунок зовнішнього джерела. Запасена в продуктах фотосинтезу енергія - основне джерело енергії для тварин та людей.
Щорічно в результаті фотосинтезу на Землі утворюється 150 млрд. тонн органічної речовини і виділяється близько 200 млн. тонн вільного кисню.
Кругообіг кисню, вуглецю та інших елементів, що беруть участь у фотосинтезі, підтримує сучасний склад атмосфери, необхідний для життя на Землі. Фотосинтез перешкоджає збільшенню концентрації СО2, запобігаючи перегріву Землі внаслідок так званого "парникового ефекту".
Оскільки зелені рослини являють собою безпосередню або опосередковану базу харчування всіх інших гетеротрофних організмів, фотосинтез задовольняє потребу в їжі всього живого на нашій планеті. Він - найважливіша основа сільського і лісового господарства. Хоча можливості впливу на нього ще не великі, але все ж і вони, в якійсь мірі використовуються. При підвищенні концентрації вуглекислого газу в повітрі до 0,1% (проти 0,3% в природній атмосфері) вдалося, наприклад, підвищити врожайність огірків і томатів втричі.
Квадратний метр поверхні листя протягом однієї години продукує близько одного грама цукру, це значить, що всі рослини, за приблизною оцінкою, вилучають з атмосфери від 100 до 200 млрд. тонн С на рік. Близько 60% цієї кількості поглинають ліси, що займають 30% непокритою льодами поверхні суші, 32% – окультурені землі, а решта 8% - рослини степів і пустельних місць, а також міст і селищ.
ОБМІН ЖИРІВ
Енергетичний обмін жирів
Жирні кислоти в клітинах гетеротрофів транспортуються в мітохондрії за допомогою специфічного переносника, де піддаються окисненню з утворенням ацетил-КоА, який надходить у цикл Кребса (у разі окиснення 1 молекули пальмітинової кислоти до CO2 і води утворюється 140 молекул АТФ). Цикл Кребса, таким чином, є універсальним клітинним циклом, який сполучає між собою обмін вуглеводів і жирів. Процес розщеплення жирних кислот може відбуватися тільки в аеробних умовах.
Пластичний обмін жирів
Ацетил-КоА може правити за джерело для синтезу жирних кислот і холестеролу (а також стероїдних гормонів у відповідних клітинах ендокринних залоз). У жировій тканині жирні кислоти та глюкоза перетворюються на тріацилгліцероли і запасаються у формі ліпідних (жирових) крапель доти, доки не будуть затребувані іншими органами. У разі голодування тріацилгліцероли жирової тканини хребетних і деяких безхребетних тварин розщеплюються до жирних кислот і гліцеролу, які виходять у кров і слугують джерелом енергії для міокарду (у клітинах печінки гліцерол також здатний перетворюватися на глюкозу). Тріацилгліцероли можуть частково синтезуватися з глюкози у разі її окиснення до ацетил-КоА.
ОБМІН БІЛКІВ
Пластичний обмін – синтез білка
Амінокислоти, що утворюються внаслідок розщеплення харчового білка гетеротрофами, транспортуються в цитоплазму клітин. Далі вони включаються в синтез клітинних білків. Замінні амінокислоти можуть утворюватися в клітині з різних речовин.
Незамінні амінокислоти обов'язково мають надходити в гетеротрофні клітини з навколишнього середовища. Для різних організмів незамінними є різні амінокислоти.
Клітини автотрофів здатні самостійно синтезувати всі необхідні амінокислоти, використовуючи нітратні сполуки, поглинені з ґрунту. При цьому витрачається енергія, акумульована в хімічних зв'язках АТФ у процесі фотосинтезу.
Процес синтезу білка складається з декількох етапів:
1) транскрипція іРНК на матриці ДНК;
2) транспорт іРНК через ядерні пори в цитоплазму клітини;
3) формування комплексу іРНК з великою і малою субодиницями рибосом;
4) трансляція – процес реалізації інформації, закодованої в нуклеотидній послідовності ІРНК, в амінокислотну послідовність білка;
5) посттрансляційні модифікації синтезованої білкової молекули.
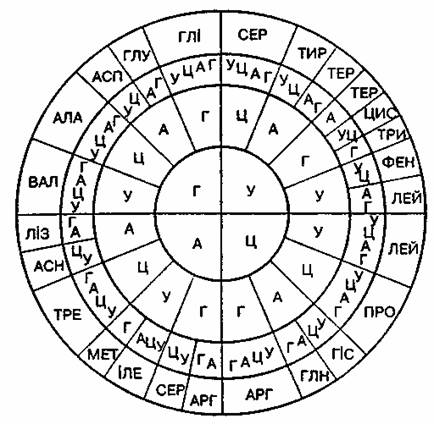
Генетичний код – властива всім живим організмам єдина система збереження спадкової інформації в молекулах нуклеїнових кислот у вигляді послідовності нуклеотидів.
Властивості генетичного коду
Триплетний – кожній амінокислоті відповідає трійка нуклеотидів ДНК (РНК) – кодон
однозначний – один триплет кодує лише одну амінокислоту
вироджений – одну амінокислоту можуть кодувати декілька різних триплетів;
універсальний – єдиний для всіх організмів, які існують на Землі;
не перекривається – кодони зчитуються один за одним, з однієї певної точки в одному напрямку (один нуклеотид не може входити одночасно до складу двох сусідніх триплетів);
між генами існують «розділові знаки» – ділянки, які не несуть генетичної інформації, а лише відокремлюють одні гени від інших. Їх називають спейсерами
Реакції матричного синтезу.
Матриця – це зразок, на якому можна створити певну кількість копій.
Матричний синтез – специфічна особливість молекулярних процесів живих організмів здійснювати біосинтез на матрицях.
Реакціями матричного синтезу є: реплікація ДНК, транскрипція та трансляція. У процесі реплікації ДНК матрицями виступають лідируючий та відстаючий ланцюги материнської ДНК, з якої за принципом коплементарності та за напівконсервативним шляхом утворюються дві ідентичні дочірні ДНК. У процесі транскрипції матрицею виступає один з ланцюгів ДНК, з якою за принципом комплементарності синтезується незріла про-іРНК.
Етапи біосинтезу білка
|
Етап |
Місце |
Процеси |
|
Транскрипція |
Каріоплазма |
Фермент РНК-полімераза розщеплює подвійний ланцюг ДНК і на одному з ланцюгів за принципом комплементарності синтезує молекулу про-іРНК. За допомогою спеціальних ферментів про-іРНК перетворюється в активну форму іРНК, яка надходить з ядра до цитоплазми клітини |
|
Активація амінокислот |
Цитоплазма |
Приєднання амінокислот за допомогою ковалентного зв'язку до певної тРНК. тРНК транспортує амінокислоти до місця синтезу білка |
|
Трансляція |
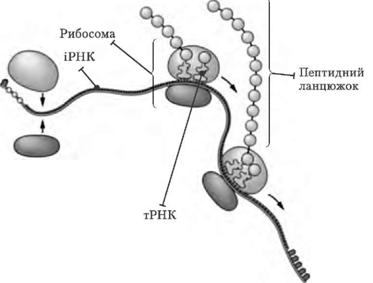
Рибосоми |
Під час синтезу білка рибосома насувається на ниткоподібну молекулу іРНК таким чином, що іРНК опиняється між її двома субодиницями. У рибосомі є особлива ділянка – функціональний центр. Його розміри відповідають довжині двох триплетів, тому в ньому водночас перебувають два сусідні триплети іРНК. В одній частині функціонального центру антикодон тРНК пізнає кодон іРНК, а в іншій – амінокислота звільнюється від тРНК. Коли рибосома досягає стоп-кодону, синтез білкової молекули завершується |
|
Утворення природної структури білка |
Ендоплазматична сітка |
Білок набуває певної просторової конфігурації. За участю ферментів відбувається відщеплення зайвих амінокислотних залишків, введення фосфатних, карбоксильних та інших груп тощо. Після цих процесів білок стає функціонально активним |
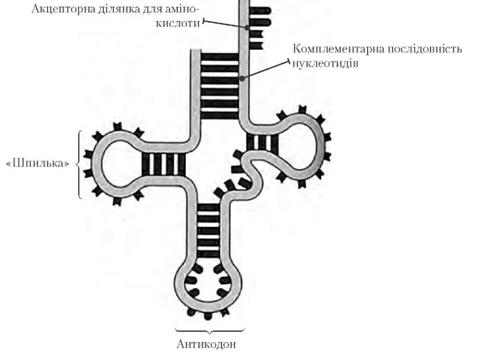
Транспорт амінокислот до рибосом забезпечують тРНК. Для кожної амінокислоти існує специфічна тРНК. Молекули тРНК містять близько 80 нуклеотидів. Усі молекули мають подібну структуру: у кожної є акцепторна ділянка, до якої приєднується відповідна амінокислота, ділянка, що містить антикодон – послідовність з трьох нуклеотидів, комплементарну кодону іРНК, який відповідає певній амінокислоті.
Транспортна PHK з приєднаною до неї амінокислотою надходить до рибосоми та зв'язується антикодоном із комплементарним триплетом (кодоном) молекули ІРНК. Зв'язування відбувається в чітко визначеному місці – на так званій А-ділянці рибосоми. У цей момент на P-ділянці (вона розташована поряд з А-ділянкою) вже є тРНК, яка утримує кінець поліпептидного ланцюга, який росте. Амінокислота, закріплена на тРНК, яка щойно надійшла, утворює пептидний зв'язок з СООН-кінцевою амінокислотою поліпептидного ланцюга, і тРНК, яка до цього була розташована на P-ділянці, відділяється від рибосоми і здатна транспортувати іншу таку ж амінокислоту. Це приводить до переміщення тРНК, що залишилася (з якою тепер зв'язані амінокислоти синтезованого білка), на P-ділянку, що звільнилася. Тепер А-ділянка доступна для прикріплення наступної молекули тРНК, антикодон якої комплементарний кодону іРНК (остання також перемістилася відносно А-ділянки на один триплет). Так триває доти, доки в А-ділянці рибосоми не опиниться кодон іРНК, який не кодує жодної амінокислоти – стоп-кодон. В еукаріотів стоп-кодонами є триплети УАА, УАГ і УГА. Для них не існує комплементарного антикодону тРНК; відсутність тРНК в А-ділянці викликає відщеплення поліпептидного ланцюга від тРНК, що знаходиться в Р-ділянці. Трансляція припиняється.
Для більшості клітин синтез білка – найбільш енергоємний з усіх біосинтетичних процесів. Утворення кожного нового пептидного зв'язку супроводжується розщепленням щонайменше 4 молекул АТФ: 2 з них витрачаються на приєднання амінокислоти до відповідної тРНК, а ще 2 – на зв'язок тРНК з А-ділянкою і пересування рибосоми вздовж ланцюга ІРНК.
Збирання одного білка триває в середньому від 20 до 560 секунд. Але навіть ця величезна швидкість може бути збільшена, якщо синтез поліпептидного ланцюга відбувається на полірибосомальному комплексі (полісомі). В останньому випадку нова рибосома приєднується до молекули іРНК відразу ж після того, як попередня зв'яже між собою достатню кількість амінокислот, щоб звільнити їй місце (це відповідає приблизно 80 нуклеотидам).
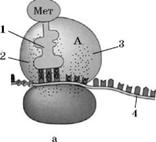
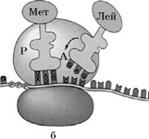
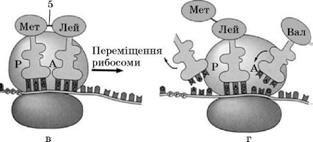
Синтез білка:
1 – тРНК, 2 – Р-ділянка рибосоми, 3 – А-ділянка рибосоми,
4 – ІРНК, 5 – ковалентний зв'язок
Посттрансляційні модифікації.
Після закінчення трансляції до утвореної молекули білка можуть приєднуватися різні органічні молекули – вуглеводи, жирні кислоти тощо. Цей процес відбувається в ендоплазматичному ретикулумі й апараті Гольджі та називається посттрансля- ційною модифікацією.