ОСНОВИ ЕВОЛЮЦІЙНОГО ВЧЕННЯ
Основи еволюційного вчення
| Сайт: | Підготовка до ЗНО - Освітній портал "Академія" |
| Курс: | Підготовка до ЗНО з біології |
| Книга: | ОСНОВИ ЕВОЛЮЦІЙНОГО ВЧЕННЯ |
| Надруковано: | Гість |
| Дата: | Sunday 1 June 2025 1:12 PM |
1. Основи еволюційного вчення
ОСНОВИ ЕВОЛЮЦІЙНОГО ВЧЕННЯ
Еволюція (від лат. еволютіо - розгортання) - це процес необоротних змін у будові та функціях живих організмів протягом їхнього історичного розвитку. її наслідком є пристосованість організмів до умов середовища життя. Загальні закономірності, фактори, механізми і наслідки еволюції живої матерії вивчає розділ біології - еволюційне вчення.
Філогенез – це історичний розвиток як усього живого загалом, так і окремих груп (видів, родів, родин і т.д. до царств включно)/
Філогенетичний ряд – це історична послідовність предкових форм певного сучасного виду.
Становлення еволюційних поглядів
Додарвінівський період
Величезну роль у накопиченні наукових фактів відіграли Великі географічні відкриття. Період накопичення знань про різноманітні рослини і тварин увійшов у науку як описовий, інвентаризаційний період. Накопичення фактичного матеріалу висувало необхідність створення наукової термінології і системи рослин і тварин
Англійський біолог Джон Рей уперше звів вид до рангу біологічного поняття. Були встановлені три особливості виду: 1) об’єднання багатьох особин; 2) морфологічна і фізіологічна подібність між ними; 3) здатність до спільного розмноження і відтворення потомства, яке зберігає подібність до батьківських форм
К. Лінней одержав світове визнання завдяки створеній ним системі рослин і тварин. Лінней установив реальність видів, чітко акцентував репродуктивну ізоляцію між ними, виявив їхню стабільність, підготував ґрунт для постановки проблеми про їхнє походження; здійснив реформу мови ботаніки, увів наукову термінологію
Погляди Ж. Б. Ламарка. Ж. Б. Ламарк створив першу цілісну концепцію еволюції живої природи (ламаркізм). Згідно з гіпотезою Ламарка, еволюція – це процес надбання корисних ознак, які успадковуються нащадками. Види змінюються, але дуже повільно, тому непомітно. Еволюція носить прогресивний характер, тобто розвиток відбувається від простого до складного. Будь-яка мінливість спадкова й обумовлена впливом зовнішнього середовища. Рушійна сила еволюції – внутрішнє прагнення до досконалості.
Підвищення організації живих істот від нижнього ступеня до вищого в процесі еволюції Ламарк назвав градацією
Основні положення еволюційного вчення Чарлза Дарвіна
Ч. Дарвін – англійський природознавець, основоположник теорії еволюції. Під час кругосвітньої подорожі (1831 – 1836) зібрав багатий науковий матеріал, що став основою його головної праці «походження видів» (1859). Еволюція, за Ч. Дарвіном, полягає в безперервних пристосувальних змінах видів.
Основні положення вчення ч. Дарвіна
Передумова еволюції: спадкова (індивідуальна) мінливість Рушійні сили: боротьба за існування та природний добір
Еволюція – поступове ускладнення й підвищення організації живих істот (еволюція має прогресивний характер)
|
Мінливість |
||
|
Групова (неспадкова, визначена) |
Індивідуальна (спадкова, невизначена) |
Співвідносна (кореляційна) |
|
Подібна зміна всіх особин потомства в одному напрямку внаслідок впливу певних умов |
Поява різноманітних незначних відмінностей в особин одного сорту, породи, виду, якими, існуючи у схожих умовах, одна особина відрізняється від інших. Не виключена можливість і різних відхилень |
Зміна структури або функції однієї частини нерідко обумовлює також певні зміни інших |
|
Рушійні сили еволюції |
||||
|
|
|
|
|
|
|
Боротьба за існування – внутрішньовидова; – міжвидова; – з несприятливими умовами навколишнього середовища |
|
Природний добір Виживання найпристосованіших і загибель найменш пристосованих
|
||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
|
|
||||
Природний добір
Ч. Дарвін дав таке визначення природного добору: «Збереження корисних відмінностей або змін і знищення шкідливих я назвав природним добором або виживанням найбільш пристосованих». Під природним добором розуміють здійснюваний у природі процес збереження й переважного розмноження в ряді поколінь особин, що мають корисні для їхнього життя і розвитку пристосувальні ознаки, що виникли в результаті різноспрямованості індивідуальної мінливості.
|
Рушійний добір (прямий, провідний) |
Кожний вид складається не з абсолютно однакових особин. При тривалій зміні зовнішнього середовища в одному напрямку створюються умови, при яких окремі мутації виявляються корисними й зберігаються в ході добору. Зумовлює постійну зміну пристосувань видів відповідно до змін умов середовища |
|
Стабілізуючий добір |
У малозмінних умовах існування збільшується чисельність особин із середньою нормою реакції. Із покоління в покоління відтинаються крайні форми, а закріплюються організми з певною нормою реакції |
|
Дизруптивний добір (розриваючий, спрямований проти проміжних форм) |
Іноді умови зовнішнього середовища змінюються таким чином, що перевагу одержують крайні форми. Кількість таких форм швидко збільшується, що може привести до перетворення виду |
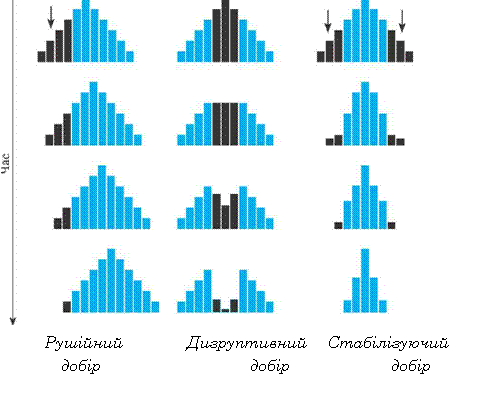
Обставини, що сприяють природному добору
1. Кількість особин і їхня різноманітність
2. Частота мутацій
3. Інтенсивність розмноження
4. Індустріальний меланізм
5. Частота зміни поколінь
6. Розміри ареалу і різноманітність умов життя в ньому
7. Ізоляція, що перешкоджає схрещуванню
Синтетична теорія еволюції сформувалася на початку 40-х рр. ХХ ст. Вона являє собою вчення про еволюцію органічного світу, розроблене на основі даних сучасної генетики, екології та класичного дарвінізму.
Основні положення синтетичної теорії еволюції
Матеріалом для еволюції служать, як правило, дуже дрібні зміни спадковості – мутації. Мутаційна мінливість – постачальник матеріалу для добору – носить випадковий і спрямований характер
Основним рушійним фактором еволюції є природний добір, що виникає на основі боротьби за існування
Найменшою одиницею еволюції є популяція
Еволюція носить дивергентний характер, тобто один таксон може стати предком кількох дочірніх таксонів, але кожний вид має єдиний предковий вид, єдину предкову популяцію
Еволюція має поступовий і тривалий характер. Видоутворення як етап еволюційного процесу являє собою послідовну зміну однієї тимчасової популяції низкою наступних тимчасових популяцій
Вид складається з безлічі підвидів і популяцій. Однак відомо чимало видів з обмеженими ареалами, у межах яких не вдається відокремити самостійні підвиди, а реліктові види можуть складатися з єдиної популяції. Доля таких видів, як правило, недовговічна
Вид існує як цілісне й замкнуте утворення. Цілісність виду підтримується міграціями особин з однієї популяції до іншої, при яких спостерігається обмін генами
Оскільки основним критерієм виду є несхрещуваність з іншими видами (репродуктивна ізоляція), то цей критерій не стосується форм, у яких не спостерігається статевий процес
Макроеволюція йде лише шляхом мікроеволюції (ті самі передумови та рушійні сили)
Будь-який реальний таксон має монофілетичне (від одного предка) походження
Еволюція має неспрямований характер, тобто не йде у напрямку якої-небудь кінцевої мети
Еволюція має незворотний характер. Організм (популяція, вид) не може повернутися до колишнього стану, вже здійсненому в низці його предків
Мікроеволюція
Мікроеволюція – це сукупність еволюційних процесів, які відбуваються на рівні популяцій. Мікроеволюція закінчується утворенням нових видів.
Відповідно до синтетичної теорії еволюції, у популяціях діють, крім основних факторів еволюції – мутацій, боротьби за існування й добору, – і так звані елементарні фактори еволюції: хвилі життя, дрейф генів, ізоляція.
Хвилі життя (популяційні хвилі) – це періодичні коливання розмірів популяцій за числом особин (С. С. Четвериков). Причини цих коливань різноманітні, вони можуть бути біотичні та абіотичні (запаси їжі, кількість хижаків, конкурентів, кліматичні умови року тощо). Після збільшення числа особин у популяції відбувається закономірне його зменшення тощо. Хвилі життя самі по собі не викликають спадкової мінливості, але вони сприяють зміні частот алелів у популяціях.
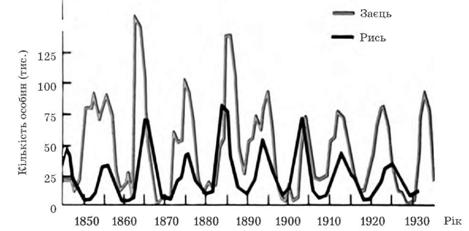
Співвідношення чисельності хижаків і жертв
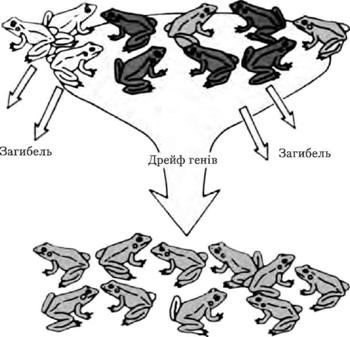
Дрейф генів відкрили С. Райт і Р. Фішер і незалежно від них М. П. Дубінін і Д. Д. Романов. Це випадкова зміна концентрації алелів у популяції. Стосується нечисленних популяцій. Випадкові події, наприклад передчасна загибель особини, що була єдиним власником якогось алеля, спричинюють до зникнення цього алеля в популяції, і навпаки – частота алеля може випадково підвищитися. Дрейф генів є чинником, який постачає матеріал для еволюції.
 |
 |
Ізоляція
Ізоляція – постійне обмеження вільного схрещування.
|
Типи ізоляції |
||||
|
Географічна |
Екологічна |
Сезонна |
Етологічна |
Генетична |
|
Пов'язана з територіальним розмежуванням груп |
Потенційні партнери по спарюванню займають різні екологічні ніші й не зустрічаються |
Терміни розмноження в потенційних партнерів настають не одночасно |
Залежить від особливостей поведінки партнерів у період розмноження |
Партнери мають істотні генетичні розходження |
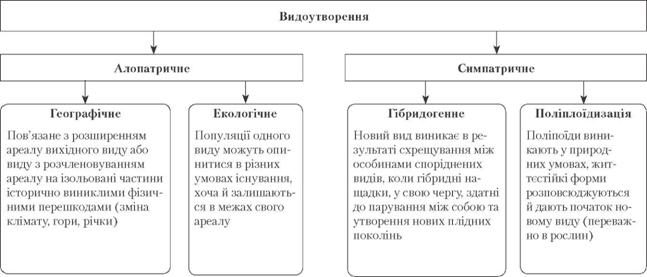
Видоутворення

Пристосованість видів не може бути абсолютною, вона завжди відносна й корисна лише в тих умовах середовища, в яких відбувався природний добір. Наприклад, форма тіла, органи дихання та інші особливості риб доцільні лише за умов життя у воді й зовсім не придатні при наземному існуванні.
Пристосування ніколи не буває абсолютно досконалим. Отруйних змій поїдають їжаки, мангусти, деякі птахи. Відносність пристосування обумовлена й тим, що саме середовище змінюється, тому адаптації, вироблені в одних умовах існування, втрачають своє значення в інших
Захисна форма тіла
Мімікрія
Застережливе забарвлення
|
|
оса
|
|
|
плями, що нагадують очі, на тілі гусені
|
|
|
гусінь схожа на суху гілку
|
|
|
Квіткова муха
|
|
|
крила комах схожі на листок
|
|
|
яскраві жовті плями на тілі саламандри
|





Макроеволюція – це надвидова еволюція, процес утворення з видів нових родів, із родів – нових родин тощо. Вона відбувається в історично тривалі проміжки часу й недоступна безпосередньому вивченню.
Способи макроеволюції
 |
Філетична еволюція
Дивергенція Конвергенція Паралелізм
Конвергентна еволюція форми тіла водних хребетних

Напрямки макроеволюції
|
Біологічний прогрес |
Біологічний регрес |
|
Зростання пристосованості організмів до навколишнього середовища (за О. М. Сєверцовим): збільшення чисельності; розширення ареалу; прогресивна диференціація – збільшення числа систематичних груп, що складають даний таксон |
Зниження пристосованості до навколишнього середовища: зниження чисельності; звуження ареалу; поступове чи швидке зменшення видового різноманіття групи може призвести до вимирання |
Шляхи досягнення біологічного прогресу
|
Ароморфоз (арогенез, морфофізіологічний прогрес) |
Ідіоадаптація (алогенез) |
Загальна дегенерація (катагенез) |
|
Підвищення організації; розвиток пристосувань широкого значення; розширення ареалу |
Розвиток пристосувань вузького значення; розширення ареалу |
Різке спрощення будови та способу життя |
Докази еволюції
Порівняння будови і процесів життєдіяльності. Завдяки дослідженням Т. Гекслі, К. Гегенбауера, О. О. Кова- левського та інших учених були розроблені еволюційні поняття про гомології, аналогії, рудименти, атавізми.
Гомології – це відповідність загального плану будови органів різних видів, зумовлені їхнім спільним походженням
Аналогії – це зовнішня подібність видів за будовою органів, які мають різне походження, проте виконують однакові функції
Рудименти – це органи, недорозвинені чи спрощені у певних видів порівняно з подібними утворами предкових форм унаслідок втрати своїх функцій протягом філогенезу
Атавізми – прояв у окремих представників виду рис, притаманних їхнім предкам

Гомологічні органи: нога ящірки (а), крило кажана (б), плавець кита (в), рука людини (в)

Аналогічні органи: крила бабки (а), кажана (б), птаха (в)

Рудиментарні органи людини: 1 – третє повіко, 2 – м'яз, рухаючий вухо, 3 – ікла, 4 – зуби мудрості, 5 – апендикс, 6 – куприк
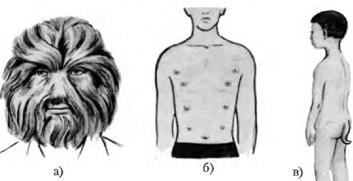
Атавізми: а) волохата людина; б) багатососковість у людини; в) хвіст у хлопчика
Відбиток на камені археоптеріксу – перехідної форми між плазунами та птахами (150 млн років тому)
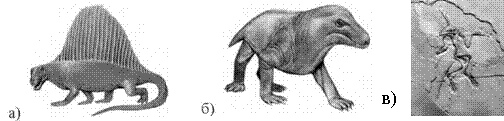
Реконструйовані перші ссавці за знайденими залишками кісток: а) диметродон, б) циногнатус (середина тріасового періоду)
 |
Вивчаються викопні залишки вимерлих організмів і виявляються їхні подібність і відмінність із сучасними організмами. В одних випадках вдалося встановити перехідні форми, в інших – філогенетичні ряди (ряди видів, що послідовно замінюють один одного).
Біогенетичний закон: онтогенез – це коротке й швидке повторення філогенезу, або кожна особина у своєму індивідуальному розвиткові (онтогенезі) повторює історію розвитку виду (філогенез), до якого вона належить (Е. Геккель, Ф. Мюллер, 1866 р.).
М. О. Сєверцов доповнив біогенетичний закон
1) в онтогенезі зазвичай повторюється будова не дорослих стадій предків, а їхніх зародків;
2) в онтогенезі можуть виникати спеціальні пристосування до тих умов, у яких розвиваються зародки;
3) в ембріонів можуть виникати мутації, які далі змінюють ознаки дорослої тварини
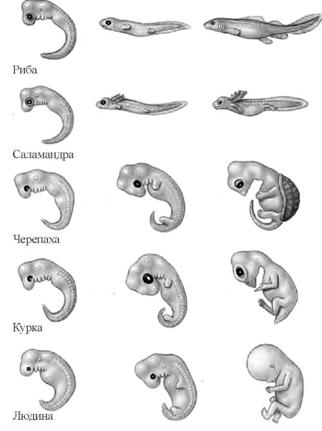
Ембріональний розвиток хребетних
Сучасні еволюційні гіпотези.
Важливим показником біологічного процвітання виду є оптимальна густота просторового розміщення особин кожної з його популяцій, яка зумовлена збалансованістю народжуваності та загальної смертності. Розглянемо найпопулярніші сучасні еволюційні гіпотези.
Неокатастрофізм (від грец. неос - новий та катастрофе - переворот) - система поглядів, яка базується на факті етапності розвитку життя на Землі, тобто гіпотеза катастроф. У 1864 р. Е. Зюсс модернізував погляди Ж. Кюв'є для пояснення швидкої заміни одних комплексів викопних організмів іншими; саме його вважають засновником неокатастрофізму.
Етапність історичного розвитку живого полягає в тому, що протягом послідовних етапів геологічної історії Землі існували певні більш-менш стабільні екосистеми зі специфічними флорою та фауною, які на межах цих етапів відносно швидко (за кілька тисяч років) замінювались іншими. Об'єктивні підтвердження цього явища дає вивчення темпів вимирання одних і появи інших систематичних груп організмів різного рангу. Протягом певного, досить тривалого часу (мільйони та десятки мільйонів років) незначне вимирання одних груп врівноважується появою екологічно близьких їм інших; на межах періодів та окремих епох (епоха - геологічний підрозділ всередині періоду, наприклад рання, або нижня, середня, пізня, або верхня, крейда - крейдяний період) незбалансовані вимирання зареєстровані наприкінці попереднього, а незбалансоване швидке зростання видового різноманіття - на початку наступного періоду чи епохи.
Наприклад, наприкінці середньокрейдяної епохи відбулося масове вимирання багатьох груп голонасінних, комах, динозаврів (зникло 5 родин з 11), ссавців та значного числа видів інших організмів. Навпаки, на початку другої половини крейдяного періоду відбувалось бурхливе видоутворення: виникають і зазнають значної адаптивної радіації покритонасінні та комахи-запилювачі (мухи, деякі групи жуків, бджоли, денні метелики тощо), з'являються 10 нових родин динозаврів, кілька рядів справжніх птахів, сумчастих і плацентарних ссавців. Протягом пізньокрейдяної епохи число цих новоутворених груп стабілізується, а в її кінці знову спостерігається незбалансоване вимирання, зокрема динозаврів, яке замінюється бурхливим видоутворенням на початку наступного періоду (палеоген). Вимирання динозаврів стало підґрунтям для різних фантастичних пояснень його причин. Так, однією з популярних гіпотез вимирання динозаврів є їхня раптова загибель внаслідок зіткнення Землі з астероїдом. Однак жодних фактичних підтверджень цього не знайдено.
Отже, на межах геологічних періодів та епох відбуваються різкі зміни видового складу біосфери, тобто заміна одних біогеоценозів іншими. Причини цього явища вчені вбачають у неперіодичній зміні інтенсивності екологічних факторів, яка перевищує межі витривалості біогеоценозів, руйнуючи їх, що і спричинює масове вимирання спеціалізованих видів. Неспеціалізовані види як екологічно пластичні опановують простір зруйнованих екосистем і формують там нові стійкі біогеоценози, адаптуючись до їхніх умов. Цим пояснюють бурхливу адаптивну радіацію на початку періодів та епох.
У житті біосфери геологічні катастрофи спричинюють загальні (глобальні) або місцеві (локальні), тобто в межах певної території, біоценотичні кризи. Вони можуть бути спричинені не лише геологічними, але й біогенними причинами. Наприклад кризу середини крейдяного періоду спричинила поява покритонасінних рослин; сучасна екологічна криза внаслідок дії антропогенного фактору, що може катастрофічно закінчитись.
Гіпотеза нейтральності молекулярної еволюції.
Гіпотезу нейтральності молекулярної еволюції вперше опублікував японський генетик Мотоо Кімура у 1968 р. і розвинув у багатьох своїх наступних працях. Незалежно від М. Кімури американські біохіміки Д. Кінг та Т. Джукс дійшли майже тих самих висновків про нейтральну еволюцію (1969).
М. Кімура зі співробітниками провели низку експериментів над дрозофілами та математично обробили результати, а також вивчили генофонди природних популяцій різних видів. Виявилось, що в будь-якій природній чи штучній популяції 7 – 15 % організмів були гетерозиготні по кожному з локусів. Від 30 до 50 % генів були представлені декількома алелями. Це визначає поліморфізм білків, тобто явище, коли кожний білок у різних організмів одного виду фактично перебуває в кількох генетично зумовлених формах. Сумарна кількість алелів, які несуть інформацію про поліморфні білки, у популяціях становить багато тисяч, тому число різних поліморфних білків сягає мільйонів і більше. М. Кімура припустив, що більшість цих білків нейтральна, тобто не шкідлива і не корисна для організму і не підлягає дії добору. Пізніше було встановлено, що найпоширеніші мутації в структурі ДНК або не порушують послідовності амінокислот при синтезі поліпептидів, або відбуваються в ділянках, які не кодують інформацію про амінокислоти. Таким чином, у мікроеволюції чільне місце належить випадковим процесам (мутації, дрейф генів, елімінація), які змінюють частоту зустрічальностей алелів, а не природному добору чи боротьбі за існування.
Гіпотеза сальтаціонізму.
Сальтаціонізм (від лат. сальто - стрибаю) - система поглядів на темпи еволюції як стрибкоподібні зміни, що відбуваються за незначний проміжок часу і спричинюють виникнення нових видів, родів тощо. Ці погляди побудовано на уявленнях про те, що темпи еволюції зумовлені швидкістю змін умов довкілля. Зрозуміло, що кожний вид має бути адаптованим до умов середовища життя в будь-який момент свого існування. Якщо ж він не встигає пристосуватися до змін довкілля, то вимирає. Тому в стабільні періоди темпи еволюції незначні або ж вона не відбувається зовсім. Під час біоценотичних і біосферних криз ці темпи зростають, і видоутворення в масштабах геологічної історії Землі відбувається практично миттєво, стрибкоподібно.
Основні положення гіпотези перерваної рівноваги.
Відомо, що біологічні системи всіх рівнів організації здатні функціонувати лише за сталого або періодично змінного середовища. Неперіодичні зміни інтенсивності екологічних факторів, що перевищують межі стійкості живих систем, призводять до їхнього руйнування. Найвразливіші до таких впливів біогеоценози та біосфера в цілому. На цьому ґрунтується еволюційна гіпотеза перерваної рівноваги, запропонована американськими вченими Н. Елреджем, С. Гоулдом та С. Стенлі в 70-х - 80-х роках XX сторіччя та значно доповнена дослідженнями російських ботаніків та екологів С.М. Розумовського і В.А. Красилова. Її основні положення такі:
1. Кожен етап історичного розвитку біосфери має відносно стабільні кліматичні умови і характеризується певними біогеоценозами з притаманними їм спеціалізованими екологічно непластичними видами (ценофілами). Тобто існує динамічна рівновага між біотичними та абіотичними складовими екосистем і біосфери в цілому.
2. За різких (катастрофічних) змін довкілля, спричинених абіотичними, біотичними чи антропогенними факторами, необоротно порушується рівновага в екосистемах: настають локальні чи біосферні екологічні (біогеоценотичні) кризи.
3. Під час біогеоценотичних криз внаслідок руйнування стабільних екосистем вимирають ценофіли. Виживають лише ті групи (переважно мало спеціалізовані екологічно пластичні види - ценофоби), які внаслідок притаманної їм екологічної пластичності встигають адаптуватись до швидких змін довкілля.
4. Під час стабілізації умов довкілля відновлюється рівновага в біосфері внаслідок зміни самих екосистем та їх населення відповідно до нових умов. У цей час відбувається швидка адаптивна радіація груп організмів, що пережили кризу, завдяки якій виникають нові стабільні біогеоценози з новими ценофільними видами.
5. Темпи еволюції нерівномірні: вони мають характер повільних незначних змін або зовсім непомітні в стабільних біогеоценозах та багатократно прискорюються під час екологічних криз.
Гіпотези адаптивного компромісу.
Сучасний російський палеонтолог та зоолог О.П. Расніцин наприкінці 80-х років XX сторіччя звернув увагу на відоме явище відносної пристосованості організмів до середовища життя. Відомо, що вид у формі популяцій одночасно пристосовується до кожного з факторів та до їхнього комплексного впливу.
Зміцнення зовнішніх захисних утворів ( розглядаємо мал. 40.4 с. 243) знижує рухливість видів; збільшення розмірів і маси тіла тварин зменшує їхню відносну тепловіддачу, однак веде до необхідності споживання великої кількості поживних речовин, тому такі види дуже вразливі щодо нестачі їжі.
Отже, вузька пристосованість видів до певних умов довкілля (спеціалізація) забезпечує максимальне використання ресурсів середовища життя, але водночас знижує здатність адаптації до нових умов (розглядаємо мал. 40.5 с. 243).
Таким чином, адаптивний компроміс - це можливість пристосування виду до всього комплексу умов довкілля лише за рахунок неповних адаптацій до дії окремих чинників.
Наука - це система знань про матеріальні та нематеріальні явища, здобутих різними методами. Серед науковців було і є багато і віруючих, і атеїстів; однак їхні релігійні уподобання не впливали і не впливають на планування, проведення та пояснення результатів досліджень. Так, глибоко віруючими були біологи К. Лінней, Ж. Кюв'є, Ч. Дарвін, Г. Мендель, геолог Ч. Лайєль та багато інших; не менше також учених-атеїстів, починаючи з Ж.-Б. Ламарка.
Релігія в Україні, як відомо, відокремлена від держави, у чому і полягає конституційне забезпечення свободи совісті. Ніхто з державних службовців не має права діяти в інтересах якоїсь релігії чи, навпаки, вести атеїстичну пропаганду.