ІНДИВІДУАЛЬНИЙ РОЗВИТОК ОРГАНІЗМІВ
1. Індивідуальний розвиток організмів
ІНДИВІДУАЛЬНИЙ РОЗВИТОК.
Індивідуальний розвиток організму, уся сукупність його перетворень від зародження (запліднення яйцеклітини або поділу материнської клітини) до кінця життя, називається онтогенезом. Цей термін був запроваджений німецьким природодослідником Е. Геккелем у 1866 р.
В одноклітинних організмів онтогенезом вважається клітинний цикл – період життя клітини від моменту її утворення до моменту закінчення акту поділу. В онтогенезі багатоклітинних організмів виділяють такі основні періоди (етапи):
1) ембріональний – розвиток до виходу організму із зародкових оболонок;
2) постембріональний – до досягнення статевої зрілості;
3) дорослий стан, включаючи старіння і смерть.
Відповідно до іншої класифікації виділяють такі етапи онтогенезу:
1) передзародковий – включає розвиток статевих клітин і запліднення;
2) ембріональний;
3) постембріональний;
4) репродуктивний – період, коли організм здатний розмножуватися;
5) пострепродуктивний – втрата репродуктивної функції, старіння, смерть.
Величезну роль у становленні ембріології відіграли на початку XVII ст. поява мікроскопа та спостереження, зроблені з його допомогою голландськими дослідниками А. ван Левенгуком, Я. Сваммердамом та італійським лікарем і ембріологом Г. Мальпігі.
Характер дроблення великою мірою залежить від кількості жовтка в цитоплазмі яйцеклітини. Жовткові гранули перешкоджають просуванню веретена поділу до полюсів клітини й утворенню перетяжки, тому у тварин з великою кількістю жовтка (плазуни, птахи) дроблення неповне: ядра з відокремленими ділянками цитоплазми зосереджуються біля одного полюса клітини, а протилежний полюс заповнений жовтком. Яйцеклітини, що мають незначну кількість жовтка або не мають його взагалі (ссавці, плоскі черви), піддаються повному дробленню. Розрізняють рівномірне та нерівномірне повне дроблення. За рівномірного дроблення бластомери однакові (у ланцетника). У разі нерівномірного дроблення бластомери відрізняються за формою і розмірами (у жаби на одному полюсі зародка бластомери дрібніші, ніж на іншому).
Період дроблення закінчується формуванням бластули. У типовому випадку бластула складається із шару бластомерів, які оточують щільним кільцем порожнину – бластоцель. Бластоцель заповнена рідиною, яка за хімічним складом відрізняється від рідини зовнішнього середови У ссавців цю стадію формування зародка називають бластоцистою. Бластоциста відрізняється бластули тим, що бластомери в ній розташовуют ся в два шари: зовнішній дає початок трофобласту, а внутрішній (зародковий вузлик) – ембріоласту. На стадії бластоцисти зародок переміщається яйцепроводом до матки.
У багатоклітинних тварин під час вгинання (інвагінації) частини стінки бластули всередину бластоцелі (подібно до того, як можна натиснути пальцем на погано надутий гумовий м'яч). Зародок на цій стадії складається з двох шарів клітин (зародковиx листків): зовнішнього – ектодерми, і внутрішнього – ентодерми. Під час вгинання утворюється порожнина – гастроцель (гастральна порожнина), і отвір, яким вона сполучається з навколишнім середовищем – бластопор (первинний рот). У первинноротих тварин бластопор, розвиваючись і диференціюючись, перетворюється на рот дорослого організму, у вторинноротих – на анальний отвір. Рот у вторинноротих виникає на протилежному кінці зародка.
Бластоцель виявляється поміщеною між енто- й ектодермою. У неї проникають бластомери, що дають початок третьому зародковому листку – мезодермі. У ссавців, рептилій і птахів мезодерму утворюють клітини, які виселяються з певних зон ентодерми. У губок і кишковопорожнинних мезодерми немає, тому їх називають двошаровими тваринами.
У багатьох тварин виділяють ще одну стадію ембріонального розвитку – морулу. У морулі бластомери щільно притиснуті один до одного, а бластоцель відсутня
Гаструляція
Наступна стадія ембріонального розвитку на зивається гаструляцією, а зародок на ній стадії - гаструлою. Гаструла формується у більшості бага
Ембріональний розвиток ланцетника
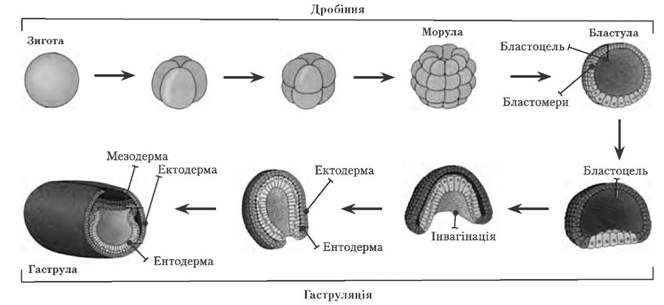
У деяких тварин гаструляція йде не шляхом інвагінації, а шляхом міграції бластомерів із стінки бластули в бластоцель. Саме такий тин утворення гаструли І. І. Мечников вважав найпримітивнішим і поклав в основу створення теорії фагоцители, згідно з якою предком сучасних багатоклітинних була примітивна тварина, яка не мала травної порожнини, клітини якої здатні були вкорінятися всередину тіла.
Для процесів клітинного поділу, які відбуваються під час гаструляції, переміщення клітин з однієї ділянки ембріона в іншу, необхідна енергія, яка утворюється внаслідок розщеплення жовтка. У рептилій, птахів, риб, багатьох комах жовток є поживним матеріалом до виходу сформованого організму або личинки з яйцевих оболонок. У ссавців та інших живородящих тварин зародок поглинає поживні речовини з організму матері, оскільки їхні яйцеклітини бідні на жовток.
Зародкові листки дають початок тканинам та органам ембріонів, які розвиваються.
|
Ектодерма |
Дає початок нервовій системі, пов'язаним з нею органам чуття, зовнішньому покриву організму, передній та задній кишкам |
|
Ентодерма |
Дає початок хорді кишечника й пов'язаним із ним органам: шлунку, печінці тощо, органам дихання – легеням |
|
Мезодерма |
Дає початок мускулатурі, усім хрящовим і кістковим елементам скелета, кровоносній і сечостатевій системам |
Процес формування органів із певних комплексів клітин ембріона називається органогенезом. Першою у процесі ембріогенезу закладається нервова система. Її розвиток – нейруляція – починається услід за гаструляцією. Ембріон на цій стадії називають нейрулою. Нейруляція у хордових починається з потовщення ділянки ектодерми (нервової пластинки) на спинному боці зародка. По краях нервової пластинки утворюються складки – нервові валики, які стають дедалі вищими, зближуються та змикаються, утворюючи нервову трубку. Канал нервової трубки перетворюється на спинномозковий канал. Головний відділ трубки виявляється ширшим; він дає початок головному мозку. Поступово в головному відділі стають помітні потовщення, які відповідають великим півкулям, виокремлюються інші відділи мозку. З випинів стінок проміжного мозку формуються закладки очей – очні пухирці.
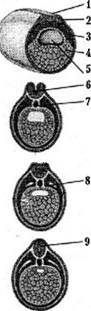
Органогенез (розвиток нервової' системи):
1. нервова пластинка,
2. ектодерма,
3. мезодерма,
4. ентодерма,
5. первинна порожнина,
6. нервовий канал,
7. хорда,
8. целом,
9. нервова грубка
Приблизно в цей же час мезодерма хордових набуває вигляду плоских клітинних шарів, розташованих з обох боків від хорди. Далі клітини мезодерми збираються у скупчення, усередині яких є порожнини. Такі посегментно розташовані клітинні маси називають сомітами. Зовнішня ділянка соміта, яка прилягає до ектодерми, є джерелом розвитку дерми, середня йде на побудову поперечно-смугастої мускулатури, а внутрішня розвивається в хрящову та кісткову тканини. Певні ділянки мезодерми диференціюються в гладеньку мускулатуру кишечнику та кровоносних судин. Клітини мезодерми, які лежать поряд із сомітами, є джерелом формування видільної системи.
Зародкові оболонки
Крім формування органів зародкові листки утворюють у рептилій, птахів і ссавців зародкові оболонки – амніон, хоріон, алантоїс. Перелічені класи об'єднують у групу амніот, яка не має систематичного рангу. Зародкові оболонки, які формують навколо зародка вологе середовище, відіграли важливу роль у відокремленні процесів розмноження амніот від водного середовища.
Амніон розвивається зі складок ектодерми та мезодерми, які підіймаються і замикаються над зародком. Амніотична порожнина, що утворюється при цьому, заповнюється амніотичною рідиною і захищає зародок від механічних ушкоджень.
Хоріон плацентарних ссавців (у плазунів і птахів цю оболонку зазвичай називають серозною) закладається ще на стадії бластоцисти, утворюючись з клітин трофобласта і мезодерми. Він безпосередньо контактує зі стінкою матки своїми ворсинками.
Загальні особливості ембріогенезу відображені в біогенетичному законі Е. Геккеля: «Онтогенез є коротке та швидке повторення філогенезу»
Алантоїс формується як виріст заднього відділу кишки зародка; у його утворенні беруть участь ентодерма та мезодерма. У рептилій і птахів алантоїс виконує дві важливі функції.
По-перше, він працює як зародковий орган виділення, накопичуючи продукти метаболізму. По-друге, алантоїс править за орган дихання ембріона: у ньому розвивається густа сітка кровоносних судин, що прилягає до яєчної шкаралупи, проникної для кисню. У ссавців у зв'язку з внутрішньоутробним розвитком плоду функція алантоїса частково втрачається. Його судини проникають у ворсинки хоріона, формуючи судини пуповини
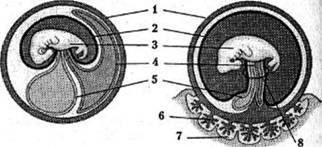
Зародкові оболонки:
1. - хоріон,
2. - амніон,
3. - ембріон,
4. - алантоїс,
5. - жовтковий мішок,
6. - зародкова ділянка плаценти,
7. - материнська ділянка плаценти,
8. - пуповина.
Запас жовтка зародків хордових зосереджується в жовтковому мішку – вирості кишечнику, стінки якого утворюються з ентодерми. Жовтковий мішок рептилій і птахів добре розвинений, тоді як у ссавців він практично не містить жовтка. Його виникнення в останніх можна вважати яскравим прикладом прояву рис розвитку еволюційних предків.
О. О. Ковалевський вперше описав гаструлу, назвав її «кишковою личинкою».
Е. Геккель запропонував назву «гаструла».
К. Бер перший установив, що під час інвагінації утворюються зародкові листки.
ПОСТЕМБРІОНАЛЬНИЙ РОЗВИТОК ОРГАНІЗМІВ
Способи відтворення
У тваринному світі відомо три способи відтворення потомства: яйцеродіния, яйцеживородіння і живородіння.
Я й ц е р о д і н н я м називається такий спосіб відтворення, за якого розвиток зародка відбувається поза тілом самки – у зовнішньому середовищі, під захистом яйцевих оболонок. Яйцеродіния характерне для більшості безхребетних, круглоротих, риб, амфібій, рептилій, птахів і деяких ссавців.
У разі яйцеживородіння зародок розвивається в тілі матері, проте не отримує від неї поживних речовин. Розвиток відбувається за рахунок запасних речовин яйцевих оболонок (деякі кліщі, риби, плазуни).
У разі ж и в о р од і н н я зародок розвивається в материнському організмі, харчується безпосередньо від нього за допомогою спеціальних пристосувань (виростів жовткового мішка тощо). Живородіння характерне для деяких червів, членистоногих, молюсків, багатьох акул, черепах, деяких ящірок, змій, переважної більшості ссавців. У плацентарних ссавців взаємодія зародка з материнським організмом досягла найбільшої складності. У цих тварин з хоріона та судин алантоїса за участю слизової оболонки матки розвивається плацента – орган, через який здійснюється обмін газами та поживними речовинами між матір'ю та плодом. Тому по відношенню до плацентарних ссавців часто говорять про істинне живородіння.
Термін «живородіння» іноді вживають щодо рослин, у яких насіння проростає на материнській рослині в ще не зрілих плодах. Так, у мангрових дерев з плоду, що зав'язався, з'являється проросток із зачатком кореня. Досягнувши довжини .10 – 70 см, він обпадає і відразу ж закріплюється в мулистому грунті.
Постембріональний розвиток – період онтогенезу після народження або виходу із зародкових оболонок до настання статевої зрілості. У цей період відбуваються ріст і розвиток організму, диференціювання тканин і органів (наприклад статевих залоз у ссавців).
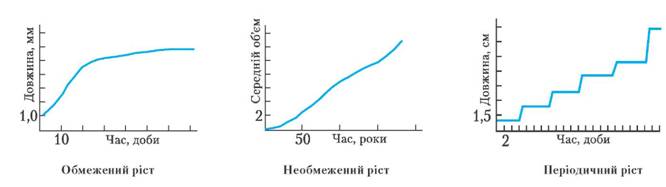
Безпосередньо після народження у більшості багатоклітинних організмів йде період росту. Ріст – збільшення розмірів і маси особини за рахунок збільшення кількості клітин та їx розтягування. Розрізняють обмежений і необмежений типи росту. У разі обмеженого типу ріст припиняється після досягнення певного віку (більшість ссавців, комах, птахи). У разі необмеженого типу росту особини ростуть упродовж усього життя (молюски, риби, рослини). Процесу росту часто властива періодичність (сезонна, добова тощо) – переривчастий ріст. Так, у рослин помірних широт ріст у зимовий час припиняється, а навесні поновлюється.
Період тимчасового фізіологічного спокою в розвитку тварин називається діапаузою. Діапауза характеризується різким зниженням інтенсивності метаболізму клітин. Вона властива, комахам, багатьом хребетним. У деяких ссавці північних широт діапауза відбувається в зимовий час (ведмеді, соні, бабаки) і називається гібернацібю (сплячкою). Діапауза комах може тривати від кількох годин до кількох років. У багатьох ракоподібних, саранових, деяких ссавців описане явище ембріональної діапаузи, за якої запліднені яйця або зародки ранніх етапів розвитку можуть тривалий час перебувати в стані спокою, не гинучи (яйця рачка Artemia salina – до 10 років).
Початок і закінчення діапаузи регулюється гормонами, а також чинниками зовнішнього середовища (температурою, довжиною світлового дня, вологістю)
Постембріональний розвиток тварин може бути прямим або супроводжуватись перетворенням – метаморфозом. У разі прямого розвитку новонароджені тварини мають основні риси організації дорослої особини та відзначаються меншими розмірами та не цілком розвиненими статевими залозами. Постембріональний розвиток зводиться до росту та досягнення статевозрілості (прісноводна гідра, багато нематод, більшість хребетних).
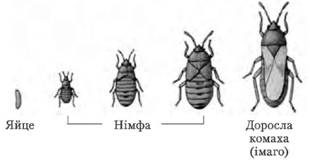
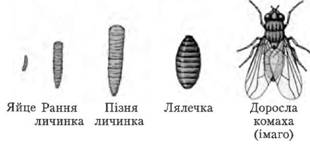
Глибоке перетворення будови організму, у процесі якого личинка перетворюється на дорослу особину, називається метаморфозом. У багатьох комах (бабки, таргани, богомоли, терміти) личинка подібна до дорослої комахи; зміни в організації супроводжуються в основному поступовим розвитком крил. У цьому випадку говорять про розвиток з неповним перетворенням. У більшості комах (жуки, лускокрилі, перетинчастокрилі) личинка червоподібна та не подібна до імаго ні зовнішнім виглядом, ні внутрішньою будовою, ні способом живлення. Тому перехід від личинкової стадії до імаго здійснюється через стадію лялечки. У цьому випадку говорять про розвиток із повним перетворенням.
Розвиток із перетворенням має ряд переваг перед прямим способом розвитку.
1. Активне харчування та ріст
2. Раціональне використання ресурсів
3. Розселення
4. Забезпечення зараження хазяїв (паразити)
Личинка, як правило, використовує джерело їжі, відмінне від дорослої особини, що приводить до послаблення внутрішньовидової конкуренції за ресурси.
Метаморфоз є і в деяких рослин. Так, у багаторічних трав відбувається відмирання надземного пагона та перехід у кореневище або бульбу на час несприятливого періоду
У губок, кишковопорожнинних, багатощетинкових червів личинка рухома і слугує для розселення виду.
Життєвий цикл – період між однаковими фазами розвитку двох або більшої кількості наступних поколінь.
|
Життєвий цикл |
|
|
Простий |
Складний |
|
Усі наступні покоління не відрізняються один від одного. Немає чіткого чергування поколінь |
Супроводжується закономірним чергуванням поколінь: статевого і нестатевого |
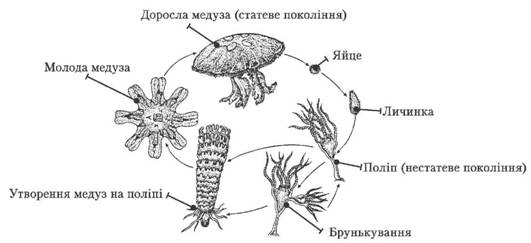
Життєвий цикл сцифоїдних
Життєвий цикл рослин
Спорофіт – нестатеве диплоїдне покоління рослин – рослина або частина рослини, де дозрівають спори.
Спорангії – орган, у якому утворюються спори.
Гаметофіт – статеве гаплоїдне покоління рослин – рослина або частина рослин, де дозрівають гамети.
Гаметангії – статеві органи – органи, в яких утворюються гамети.

У процесі еволюції рослин гаметофіт редукується. У покритонасінних чоловічий гаметофіт представлений пилковим зерном, жіночий – зародковим мішком, що складається із 7 клітин.
СТАРІННЯ І СМЕРТЬ ОРГАНІЗМІВ
Старіння – закономірний процес вікових змін, що веде до зниження адаптаційних можливосте' організму, збільшення ймовірності смерті. Старіння властиве всім організмам і перебігає на молекулярно-генетичному, клітинному, тканинному, органному рівнях організації живого.
Смерть – припинення життєдіяльності організму. Основним біологічним сенсом смерті можна вважати підтримання колообігу речовин та енергії в біосфері.
ЖИТТЄВИЙ ЦИКЛ
Життєвий цикл – це період між однаковими фазами розвитку двох або більшої кількості послідовних поколінь. У багатоклітинних організмів індивідуальний розвиток завершується природною смертю. Безперервність життєвого циклу організмів забезпечують гамети (статеві клітини), які передають спадкову інформацію організмам дочірнього покоління.
Розрізняють прості та складні життєві цикли.
За простого життєвого циклу всі покоління не відрізняються одне від одного. Прості життєві цикли характерні для різних тварин: гідри, молочнобілої планарії, дощового черв'яка, річкового рака, павука-хрестовика, плазунів, птахів, ссавців.
Складні життєві цикли супроводжуються закономірним чергуванням різних поколінь або складними перетвореннями організму під час розвитку. Так, у деяких водоростей (бурих, червоних) чергується статеве покоління, переважно гаплоїдне, з нестатевим, переважно диплоїдним. Серед вищих рослин лише у мохоподібних переважає статеве покоління, тимчасом як у інших (папоротеподібні, хвощеподібні, плауноподібні, голонасінні, покритонасінні) – нестатеве.
Складні життєві цикли, які супроводжуються зміною різних поколінь спостерігають у різних групах тварин. Так, у життєвому циклі багатьох найпростіших (форамініфери, споровики) і кишковопорожнинних відбувається закономірне чергування поколінь, які розмножуються статевим і нестатевим способами.
Нестатеве покоління цієї тварини – поліпи – розмножуеться брунькуванням, утворюючи нові поліпи. За допомогою поперечного поділу поліпи дають початок особинам статевого покоління - медузам. Чоловічі й жіночі особини медуз розмножуються статевим способом. Із заплідненої яйцеклітини розвивається личинка, яка деякий час плаває за допомогою війок, а згодом осідає на дно і перетворюється на поліп.
У інших тварин (наприклад, представників плоских червів – сисунів, у деяких членистоногих - попелиць, дафній) у життєвому циклі чергуються покоління, які розмножуються статевим способом і партеногенетично.
Наприклад, самки дрібних рачків дафній тривалий час розмножуються партеногенетично, відкладаючи незапліднені яйця. З них виходить наступне покоління самок, які знову відкладають незапліднені яйця. Але за певних змін умов довкілля (зниження температури, нестача їжі, підвищення солоності води тощо) з незашгіднених яєць виходять не тільки самки, а й самці. У самок формуються яйця, розвиток яких можливий лише після запліднення.
Ці яйця містять значні запаси поживних речовин (жовток), після запліднення вкриваються щільною оболонкою і здатні переживати періоди несприятливих умов. З настанням сприятливих умов із них виходить нове покоління самок, які розмножуються партеногенетично.
Подібний тип життєвого циклу спостерігають і у комах – попелиць.
Чергування поколінь, які розмножуються статевим способом і партеногенетично, має важливе біологічне значення для тих організмів, які мешкають у мінливих умовах довкілля і не можуть переживати несприятливі періоди в активному стані. Статеве розмноження забезпечує безперервність існування виду, а партеногенез дає змогу повною мірою використовувати сприятливі періоди для швидкого зростання чисельності виду.
У життєвому циклі інших тварин чергуються роздільностатеве і гермафродитне покоління.
Наприклад, у одного з видів круглого черв'яка - рабдитиса - особини гермафродитного покоління паразитують у легенях жаб. Яйця, які відкладають ці особини, виводяться з організму хазяїна назовні. З них виходять личинки, з яких розвиваються особини роздільностатевого покоління. Вони мешкають у ґрунті й удвічі менші за розмірами, ніж гермафродитні. У свою чергу личинки, які виходять з яєць, відкладених особинами роздільностатевого покоління, для свого подальшого розвитку повинні потрапити в організм жаби. Там з них розвиваються особини гермафродитного покоління.
Чергування поколінь, які розмножуються різними способами (статевим і нестатевим, статевим і партеногенетично), підсилює мінливість, яка забезпечує здатність виду мешкати в різних умовах довкілля і швидко реагувати на їхні зміни.
ЕМБРІОТЕХНОЛОГІЇ ТА СТОВБУРОВІ КЛІТИНИ
Мабуть, наймолодшим напрямком сучасної медицини можна вважати клітинні технології, в яких клітини є джерелом тих або інших необхідних чинників, наприклад пухлинних антигенів під час вакцинотерапії. Але використовувати клітину можна не тільки як джерело будь-яких субстанцій, а й для регенеративноїмедицини. Тут особливий інтерес викликають технології, засновані на стовбурових клітинах. Здатність до необмеженого поділу й перетворення на різні типи клітин (так звана плюрипотентность) робить їх ідеальним матеріалом для трансплантаційних методів терапії. Найбільш доступними вважаються стовбурові клітини дорослого організму. Однак реальний потенціал їх диференціювання ще мало вивчений.
Термін «стовбурова клітина» (СК) був уведений у біологію О. О. Максимовим 1908 р. Досліджуючи процеси кровотворення, він дійшов висновку: у нашому організмі протягом усього життя зберігаються недиференційовані клітини, які можуть перетворюватися на лімфоцити та інші спеціалізовані клітини сполучної тканини й крові. Пізніше О. О. Максимов назвав ці клітини стовбуровими.
На ранніх стадіях розвитку ембріона клітини неспеціалізовані. Вони отримали назву стовбурових (СК), тому що розташовані біля основи уявного стовбура генеалогічного дерева клітин, яке вінчає корона з різних спеціалізованих клітин. На відміну від звичайних клітин, приречених виконувати чітко визначені функції в організмі, СК у процесі розвитку мають можливість набувати спеціалізації. СК розмножуються шляхом поділу, як і всі інші клітини. Відмінність полягає в тому, що вони можуть ділитися необмежено, а зрілі клітини зазвичай мають обмежену кількість циклів поділу. Тож говорять, що СК здатна до проліферації, тобто до тривалого розмноження і репродукції великої кількості клітин.
Потрапляючи в організм під час трансплантації, СК продовжують ділитися й самі знаходять місце, де їхня допомога найпотрібніша. Ця здатність СК отримала назву хоумінга. Отже, хоумінг – це здатність клітин до міграції в «потрібне місце» – «рідний» орган і тканину (у свою стовбурову нішу) або в ділянку ушкодження. Надзвичайно привабливими для використання в медицині є ембріональні стовбурові клітини (ЕСК) людини: з них можна отримувати будь-які типи клітин організму. Але багато властивостей і клітинні механізми так званої «стовбурової» клітини ставлять її дуже близько до трансформованої, ракової клітини. Саме тому так важливо сьогодні вивчати характеристики самих ембріональних клітин. За вісім років, що минули з моменту одержання перших ліній ЕСК людини, вдалося з’ясувати лише невелику частину механізмів, що забезпечують у культурі самопідтримку недиференційованих клітин або їх диференціювання.